Plant Nematology at Leeds: Anti-feedants
Proteinase inhibitors
Our strategy to improve plant defences to plant parasitic nematodes has centred on the use of proteinase inhibitors (PIs). Cysteine proteinases were selected as the main target of the defence as they represent the predominant proteinase activity in plant parasitic nematodes and are not involved in mammalian digestion. The cystatins being used are from rice and maize, so already in the human diet, and studies have shown them to be non toxic, non allergenic and readily broken down by cooking.
Protein engineering to improve the efficacy of a rice cystatin
The first cystatin to be developed was from rice. Rational design was used to develop a more potent form of the native oryzacystatin-I (Oc-I) cysteine protease inhibitor. We have used the paradigm of rational design from structural information to engineer more potent cysteine proteinase inhibitors. This approach is one that can be adapted to design improved resistance .
To assist in the design of protein engineering experiments to improve the efficacy of a cysteine proteinase inhibitor (termed a cystatin) from rice, Oryzacystatin-I (Oc-I), amino acid alignments and a molecular model of Oc-I were generated. Inhibitory activities (Ki) of wild-type and variant forms of Oc-I against the cysteine proteinase papain were measured. The (cysteine proteinase inhibitor) oryzacystatin-I (Oc-I), and several engineered Oc-I variants have been tested for efficacy in inhibiting growth and development of the plant parasitic nematode Globodera pallida. For one variant, in which residue Asp86 was deleted (Oc-IDD86), the Ki was reduced by 13- to 14- fold. When expressed in tomato hairy roots both Oc-I and Oc-IDD86 had a detrimental effect on growth and development of G. pallida. This effect was significantly greater on Oc-IDD86-expressing roots leading to a reduction in size of G. pallida females to a level at which fecundity is profoundly affected.
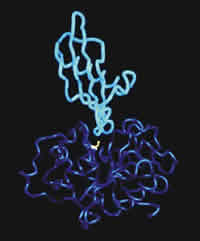
Figure: Predicted three dimensional structure of oryzacystatin-I. Oc-I (light blue) is positioned above the active site of papain (dark blue with the active cysteine shown in yellow) to show how the two molecules may interact.
Figure: Picture of tomato hairy roots
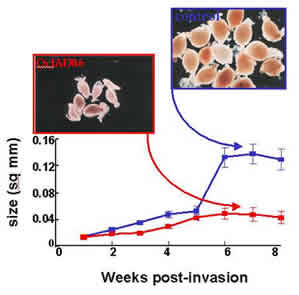
Figure. Effect of hairy roots expressing a cystatin on the growth of Globodera pallida. Red growth curve describes those animals collected from transgenic plants expressing the cystatin, the blue growth curve describes animals collected from wild type plants. Pictures show representative animals colected from the two sets of plants. The density of animals, their growth, development and fecundity were all affected.
Urwin, P.E., Atkinson, H.J., Waller,D.A. and McPherson, M.J. (1995) Engineered oryzacystatin-I expressed in transgenic hairy roots confers resistance to Globodera pallida. The Plant Journal 8: 121-131.
Further alterations by protein engineering to the amino acid sequence of Oc-I have been reported, particularly those focussed on the conserved residues of the N-terminal part of the molecule. These studies have helped define the regions of Oc-I important in inhibiting cystatins.
Urwin, P.E., Atkinson, H.J., and McPherson, M.J. (1995). Involvement of the NH2-terminal region of oryzacystatin-I in cysteine proteinase inhibition. Protein Engineering 8: 1303-1307.
Resistance to both cyst and root-knot nematodes conferred by transgenic Arabidopsis expressing a modified plant cystatin
We have tested the effects on nematode growth and development of OC-IDD86 expressed as a transgene in Arabidopsis plants. A homozygous line carrying a single copy of the OC-IDD86 gene was selected and shown by quantitative western blot analysis to be expressing OC-IDD86 as 0.4 % total soluble protein. This line was used in resistance assays with both H. schachtii and M. incognita (Urwin et al., 1996b).
On control Arabidopsis plants, most H. schachtii females became enlarged saccate in shape and were of comparable size to those recovered from cabbage plants, indicating normal growth and development. By comparison the nematodes on OC-ID D86 expressing Arabidopsis were significantly smaller. Most nematodes were fusiform and the few saccate females were all below the size of the smallest H. schachtii cyst recovered from cabbage plants. Expression of the cystatin prevented egg-laying individuals from developing, growth was arrested and some animals showed abnormal development. There was no evidence that expression of the cystatin in Arabidopsis prevented development of males of normal size.
For M. incognita the effects were similar to those observed for H. schachtii. This suggests that growth of females of both species is similarly inhibited by expression of cystatin (Urwin et al., 1996b). Three effects occur; few females reach an egg-laying size, those that do have a greatly diminished size and fecundity, and there is evidence that some saccate H. schachtii are developmentally compromised.
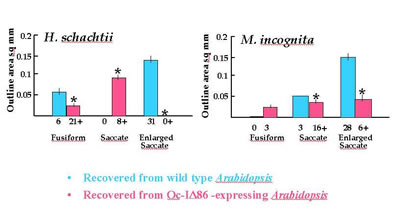
Figure. Analysis of the size of different developmental stages of A, H. schachtii and B, M. incognita taken from wild type C24 (unshaded bars) and Oc-IDD86-expressing (shaded bars) Arabidopsis at 6-8 weeks post-infection. Values for outline area are means ± SEM with n given beneath the abscissa. Statistical significant differences (P<0.05) within a category between sizes (*) and n (†)are based on t-test and c 2-test respectively.

Figure. Cysteine proteinase activity in sections of H. schachtii taken from both wild type (A) and Oc-IDD86-expressing Arabidopsis (B) viewed using fluorescence microscopy.
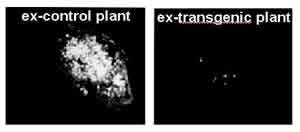
Figure. Detection of Oc-IDD86 in H. schachtii with an anti-Oc-IDD86 polyclonal antibody. From left to right, Recombinant Oc-IDD86; H. schachtii from wild type C24 Arabidopsis; H. schachtii from Oc-IDD86-expressing Arabidopsis;
Urwin, P.E., Lilley, C.J., McPherson, M.J and Atkinson, H.J. (1997) Resistance to both cyst and root-knot nematodes conferred by transgenic Arabidopsis expressing a modified plant cystatin. The Plant Journal 12: 455-461.
3. Transgenic resistance to the nematode Rotylenchulus reniformis
The level of resistance to the reniform nematode Rotylenchulus reniformis conferred on Arabidopsis thaliana by transgenic expression of proteinase inhibitors (PI) was studied over 40 days of infection. The engineered cystatin Oc-IDD86 expressed ectopically in transgenic plants as 0.4 % total soluble protein had a considerable effect on reducing parasite density and fecundity. The overall reproductive success was only ca. 20 % relative to the controls. Similar expression levels of cowpea trypsin inhibitor, a serine PI, reduced the density of the females on the plants did not reduce their fecundity. There was no evidence for additive effects when both PIs were expressed as a fusion protein using a peptide linker derived galactose oxidase. Both PIs in the fusion PI protein are known to be functional from our previous work but no effect could be determined for the CpTI component was. Supporting the observation that serine PIs are of less value than cysteine PIs for R. reniformis control. The effect of Oc-IDD86 on R. reniformis was dependent on the level of expression of different A. thaliana lines in the range of 0.1-0.4 % total soluble protein indicating that the upper level of expression is required for an effective defence. Therefore we set a threshold of at least 0.3-0.4% tsp as cystatin for selecting transgenic potato plants for field trials.
Control |
Oc-ID D86 |
CpTI |
Oc-ID D86+CpTI |
|
Female density |
1.00 ± 0.03 |
0.65 ± 0.04 * |
0.77 ± 0.03 * |
0.61 ± 0.02 * |
Mean fecundity |
1.00 ± 0.08 |
0.31 ± 0.08 * |
0.99 ± 0.06 |
0.52 ± 0.05 * |
Overall effect |
1.00 ± 0.08 |
0.19 ± 0.05 * |
0.76 ± 0.05 * |
0.31 ± 0.03 * |
Figure. Summary of female density, fecundity and overall effect, based on the product of female density and fecundity of R. reniformis parasitising PI-expressing plants relative to values for wild-type plants. Values significantly different from the wildtype using 1-way ANOVA and SNK test are indicated; * = P < 0.05.
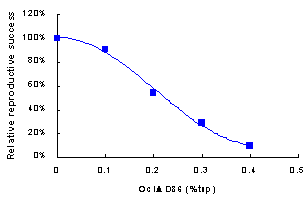
Figure: The combined effect of a reduction in fecundity of females and their lower number on reprodcutive success of Rotylenchulus reniformis on Arabidopsis thaliana over 40 days.

Figure. Rotylenchulus reniformis stained with acid fuchsin.
Summary: The work has shown that A. thaliana can act as a model host for R. reniformis. We have demonstarted that PIs produced transgenically have the potential to control R. reniformis. This approach may enable the pineapple industry to reduce its current dependence on environmentally harmful nematicides.
Urwin PE, Levesley A, McPherson MJ, Atkinson HJ. (2000). Transgenic resistance to the nematode Rotylenchulus reniformis conferred by Arabidopsis thaliana plants expressing proteinase inhibitors. Molecular Breeding 6: 257-264.